唐春梅,谢作明,程胜高
(中国地质大学(武汉)环境学院,湖北 武汉 430078)
草地植被对区域草地生态系统的稳定起着关键作用。但是,随着人口的快速增长、长期的过度放牧和全球气候变化,导致草地土壤肥力降低、植物群落结构退化和生物多样性下降。草地退化已成为我国生态环境恶化的突出问题之一。由于草地退化范围的进一步扩大及其对人类生存环境的严重影响,已引起人们的广泛关注和重视。
植物群落物种多样性是植物群落结构和功能复杂性的一种度量,受植物群落物种数量和空间分布均匀性的共同影响[1-2]。因此,植物群落物种多样性能很好地反映植物群落受环境影响的程度。到目前为止,对草地植物群落物种多样性的研究主要集中在放牧强度、浇水施肥和围栏封育等人类活动对草地群落物种多样性的影响[3-5]。其中,合理放牧可有效促进草地生态系统多样性并平衡植被动态性[6];增施有机肥和菌肥可提高土壤肥力,增加植物群落物种多样性和植物生物量[7],但过量施用化肥会引起土壤板结、生物多样性减少、草地退化[8];
短期围栏封育可显著提升植被群落功能性状和多样性指数[9],但长期围栏封育可能反而会降低植物群落物种多样性[10-11]。而在退化草地生物修复方面,主要采取人工草地建植,尽管人工草地建植可快速提高植物群落的盖度和生物量[12],但对水热条件的要求较高。而土壤藻具有抗紫外线辐射、抗高光强、抗旱、抗寒、抗贫瘠等不利环境条件的能力[13]。通过接种土壤藻培植土壤藻结皮改良退化草地,从而改善植物群落结构的研究已受到重视。土壤藻形成的生物结皮不仅可以固定沙土,防止土壤流失,而且还具有直接或者间接增加土壤营养水平、改善土壤环境、加速退化草地的生态恢复等独特的优势,土壤藻改良技术已经逐渐被人们所接受和利用[14-16]。
本文通过对土壤藻改良退化草地的土壤养分、植物群落物种组成、物种多样性和植物生产量的调查,探讨了土壤藻结皮发育对退化草地土壤肥力、植物群落结构和物种多样性的影响,为土壤藻改良退化草地提供理论依据。
1. 1 研究区概况
退化草地改良研究区位于库布齐沙漠南缘(40°21′N,109°51′E),海拔为1 040 m。该地区每年有180天风速大于5 m/s,年均降雨量为293 mm,主要集中在7—8月,且多暴雨,年均蒸发量为2 058 mm,年日照时数为3 268 h;一年中暖季短暂(5—9月),最热月份为7月份,冷季较为漫长(10—4月),最冷月份为1月份,年均气温为7.3℃,无霜期为166天[17],牧草可生长时长为174天。研究区地形地貌较复杂,主要为小型沙丘与草地相间,草地处于退化状态,沙丘以半固定沙丘为主,土壤质地以粉砂壤和沙土为主。研究区内主要的草本植物有本氏针茅(StipabungeanaTrin.)、芨芨草[Achnatherumsplendens(Trin.) Nevski]、碱蓬[Suaedacorniculata(Bunge) Bunge.]、羊草[Leymuschinensis(Trin.) Tzvel.]、披碱草(ElymusdahuricusTurcz.)、沙打旺(AstragalusadsurgensPall.)和沙蓬[Agriophyllumsquarrosum(L.) Moq.]等,同时也分布有少量的灌木,主要有柠条(CaraganaKorshinskiiKom.)、沙柳(SalixcheliophilaSchneid.)、羊柴(HedysarummongolicumTurcz Var.)和油蒿(ArtemisiaordosicaKrasch.)等。研究区在地形地貌、土壤类型、水文条件、植被特征等方面具有库布齐沙漠南缘退化草地的共同特征,具有一定的代表性。
1. 2 退化草地接种土壤藻与土壤藻结皮培植
在研究区分别选取100 m×100 m的地形地貌、土壤类型、水文条件、植被特征等环境条件大致相当的两个区域分别设定为改良区和非改良区(对照区)。
用环刀法或无菌铲在库布齐沙漠南缘退化草地采集自然藻结皮,采集厚度为5 mm,装入无菌的牛皮纸信封,封口,快速带回实验室。将藻结皮置于不同已灭菌的50 mL三角瓶中,分别加入无菌的BG11培养基,置于25℃的恒温光照培养箱中培养10 d,获得混合土壤藻的培养液。取土壤藻培养液逐级扩大培养,为研究区培植人工藻结皮提供足量混合土壤藻培养液。
为了满足试验地接种土壤藻的生长发育所需温度,并保证接种土壤藻后退化草地植被的生长,在6月中旬接种土壤藻,培植人工藻结皮。将含有土壤藻的培养液快速运到改良区,均匀接种到退化草地土壤表面,培养液中土壤藻含量大约为180 mg DW/L,1 m3土壤藻培养液大约接种100 m2退化草地。
为了保证土壤藻的成活率,在接种培养液之后的前10 d内每天用微孔喷头向退化草地浇水2次,每次浇水量与培养液接种量大致相等,并在第二个10 d内每天中午用同样方式浇水1次,保证土壤藻生长的湿润环境,随后停止浇水。改良区在培植土壤藻结皮过程中所补充的营养和水分,在非改良区也按照同样的方式补充。
1. 3 土壤样品的采集与试验前处理
在每年的6月中旬,分别从非改良区和培植土壤藻结皮1 a、2 a和3 a的改良区采集埋深为0~10 cm、10~20 cm、20~30 cm、30~50 cm的土壤。按照“S”型走向,在改良区和非改良区分别选择12处采样点,每个采样点间隔至少20 m,并在每个采样点的表层按照10 cm×10 cm大小、5 mm厚的规格采集4个样品,用于分析土壤藻结皮的生长;
同时向下挖掘土壤剖面,每个土壤层分别刮取1 kg土壤,用于分析土壤理化性质。将每4个采样点相同土壤层的样品均匀混合,设为一组,共3组,去除每组土样中的砾石、植物残枝和其他杂质,并采用四分法从各组混合土壤中保留1 kg土壤样品,分别装入采样布袋快速运回实验室。土壤样品充分混匀,分别称取少量土壤用于测定土壤含水率,剩下土壤样品在室温下自然风干,然后过100目筛,收集过筛后的土壤样品用于测定土壤养分。
1. 4 土壤藻结皮生长与土壤理化性质的测定
从采集的土壤藻结皮样品中分别称取5 g用于测定干重,5 g用于测定叶绿素含量。将5 g土壤藻结皮样品装入离心管,加入10 mL 95%的乙醇,充分摇匀,密封后置于4℃冰箱中提取24 h,取出后以4 000 r/min转速离心5 min,取上清液采用分光光度计在665 nm和649 nm波长下测定土壤藻结皮样品的光吸收值,并采用如下公式计算土壤藻结皮样品中叶绿素含量:
C=13.7×A665-5.76×A649
(1)
式中:A665和A649分别表示在665 nm和649 nm波长下样品的光吸收值;
C为样品中叶绿素含量(μg/mL)。
按照每个离心管中5 g土壤藻结皮对应10 mL乙醇,将叶绿素含量单位换算成μg/(g DS),其中DS(dry soil)表示土壤藻结皮干重。
土壤养分采用常规方法测定[18];
土壤含水率采用105℃、24 h烘干法测定;
土壤中有机质含量采用重铬酸钾-硫酸溶液氧化法测定;
土壤中全氮含量采用凯氏定氮法测定;
土壤中速效氮含量采用碱解扩散硼酸吸收法测定;
土壤中全磷含量采用高氯酸-硫酸-钼锑抗比色法测定;
土壤中速效磷含量采用碳酸氢钠浸提-钼锑抗比色法测定。本文中所有土壤重量均为土壤干重,采用烘干法测定。
1. 5 样方设置与植物群落特征调查
在植物生长最旺盛的8月份采用样方法调查植物群落结构。按照“S”型走向,分别在非改良区和改良区设置5个5 m×5 m的调查样地,每个样地间隔至少20 m,每个样地设置25个等面积的样方,并从中选取具有本样地代表性的2个样方,用于调查植物群落特征,样方面积为1 m×1 m。
分别调查样方中植物种类、植被盖度、植株密度、植被高度、植被频度和生物量,同时区分研究区内植物物种的重要性。植被高度用钢卷尺测量。样方内植物从地面刈割后,按不同植物种类称量鲜重,然后运回实验室,在实验室内自然风干,称量不同植物物种的干重[19]。植被盖度采用目测法估计。不同植物物种的重要值按照张虎等[3]的方法计算。
1. 6 植物群落物种多样性分析
植物群落物种多样性采用三类指数分析,即物种多样性指数(H)、物种丰富度指数(Ma)和物种均匀度指数(E)[20]。
物种多样性指数,采用Shannon-Wiener多样性指数,其计算公式为
(2)
物种丰富度指数,采用Margalef丰富度指数,其计算公式为
Ma=(S-1)/lnN
(3)
物种均匀度指数,采用Pielou均匀度指数,其计算公式为
E=H/lnS
(4)
式中:N表示样方内全部物种的个体总数;
Ni表示样方内第i个物种的个体数;
S表示样方内的物种数。
1. 7 数据处理与统计分析
本文采用SPSS 11.5软件对试验数据进行统计分析,对显著性水平p<0.05的试验数据在图中用误差棒标出,用Origin 8.0软件作图。
2. 1 土壤藻结皮对退化草地土壤含水率的影响
退化草地土壤藻改良过程中土壤表层叶绿素含量的变化,见表1。
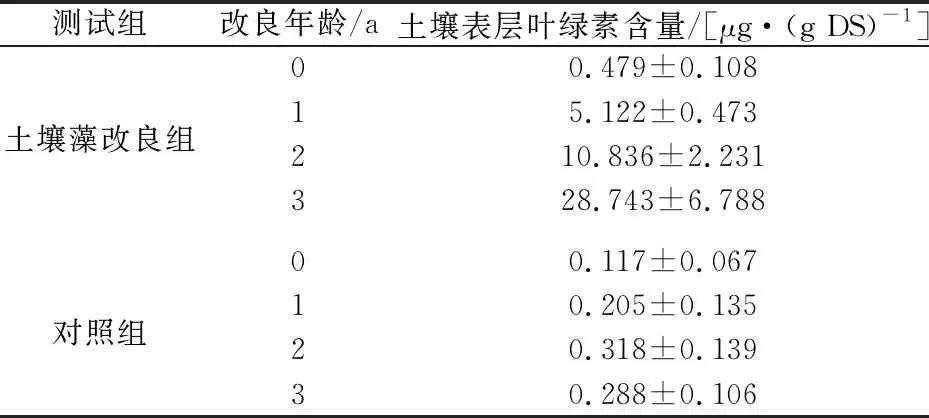
表1 退化草地土壤藻改良过程中土壤表层叶绿素含量 的变化
由表1可知:在改良区,退化草地接种土壤藻后,土壤表面生长的土壤藻叶绿素含量快速增加,经过1年、2年和3年的培养,土壤表层叶绿素含量由接种时的0.479 μg/(g DS)分别增加到5.122 μg/(g DS)、10.836 μg/(g DS)和28.743 μg/(g DS);
在非改良区的对照组,尽管没有接种土壤藻,但是由于按照改良区的管理方式补充了水分和营养,使得土壤表面的土著藻类有一定的生长,因为初始生物量极低,其生长较缓慢,土壤表层叶绿素含量增加很少,经过1年、2年和3年的培养,土壤表层叶绿素含量由接种时的0.117 μg/(g DS)分别增加到0.205 μg/(g DS)、0.318 μg/(g DS)和0.288 μg/(g DS)。
上述试验结果表明,经过1~3年的人工培植,土壤藻在土壤表层生长发育较好,形成了人工土壤藻结皮。
土壤藻结皮对退化草地土壤含水率的影响,见图1。

图1 土壤藻结皮对退化草地土壤含水率的影响
由图1可以看出:在接种土壤藻之前,退化草地土壤含水率随土壤埋深的增加而逐渐增加,且浅层土壤(0~20 cm)的含水率明显低于深层土壤(20~50 cm);
退化草地接种土壤藻之后,随着土壤藻的生长,不同埋深土壤的含水率均明显增加,其中浅层土壤(0~10 cm)的含水率增加最快,其次是埋深为10~20 cm的土壤,埋深最大的土壤含水率变化最小;
土壤藻改良3年后,埋深为0~10 cm、10~20 cm、20~30 cm和30~50 cm的土壤含水率分别增加了123.9%、63.3%、23.7%和19.9%。
上述试验结果表明,土壤藻结皮对不同埋深土壤含水率的影响差异较大,其对浅层土壤含水率的影响明显强于深层土壤。
2.2 土壤藻结皮对退化草地土壤中有机质含量的影响
土壤藻结皮对退化草地土壤中有机质含量的影响,见图2。

图2 土壤藻结皮对退化草地土壤中有机质含量的影响
由图2可以看出:退化草地经过土壤藻改良后土壤中有机质含量快速增加;
同一改良年度内,土壤中有机质含量随土壤埋深的增加而下降;
同一埋深土壤中有机质含量随土壤藻改良时间的延长而增加;
接种土壤藻后,表层土壤中有机质含量增加速度最快,其他不同埋深土壤中有机质含量也均有增加;
经过3年土壤藻结皮的发育,土壤中有机质含量是接种土壤藻前的1.2~1.3倍,而未接种土壤藻的退化草地,3年内土壤中有机质的含量几乎没有变化;
土壤藻结皮经过3年的发育,相比未接种土壤藻的退化草地土壤,埋深为0~10 cm、10~20 cm、20~30 cm和30~50 cm的土壤中有机质含量分别增加了25.7%、23.4%、19.4%和18.5%。
上述试验结果表明,在退化草地接种土壤藻形成的藻结皮可以提高不同埋深土壤中有机质的含量。
2.3 土壤藻结皮对退化草地土壤中全氮和速效氮含量的影响
土壤藻结皮对退化草地土壤中全氮和速效氮含量的影响,见图3。

图3 土壤藻结皮对退化草地土壤中全氮和速效氮含量的影响
由图3可以看出:土壤藻结皮对退化草地土壤中全氮和速效氮含量的影响与其对土壤中有机质含量的影响相同,也随土壤埋深的增加而下降,并且同一埋深土壤中全氮和速效氮含量随土壤藻结皮发育年龄的增加而增加;
土壤藻接种3年后,埋深为0~10 cm、10~20 cm、20~30 cm和30~50 cm的土壤中全氮含量分别增加了33.3%、36.4%、22.7%和31.6%,土壤中速效氮含量分别增加了52.9%、45.7%、35.0%和22.8%。由此可知,在0~30 cm埋深土壤中速效氮含量的增幅比全氮含量的增幅大,说明接种土壤藻改良退化草地不仅增加了土壤中总氮含量,而且其中可快速被植物利用的速效氮含量所占比例更大。
2.4 土壤藻结皮对退化草地土壤中全磷和速效磷含量的影响
土壤藻结皮对退化草地土壤中全磷和速效磷含量的影响,见图4。

图4 土壤藻结皮对退化草地土壤中全磷和速效磷 含量的影响
由图4可以看出:在接种土壤藻之前,0~50 cm不同埋深土壤中全磷含量的变化很小,土壤中速效磷含量的变化也几乎没有差异;
在接种土壤藻之后,同一埋深土壤中速效磷含量的变化较大,埋深为0~10 cm土壤中速效磷含量随土壤藻结皮发育年龄的增加,依次增加了13.8%、31.5%和53.7%,而埋深为10~20 cm土壤中速效磷含量依次增加了14.4%、30.8%和53.2%,埋深为20~30 cm土壤中速效磷含量依次增加了6.0%、28.1%和49.7%,埋深为30~50 cm土壤中速效磷含量依次增加了6.7%、28.4%和41.2%;
而同一埋深土壤中全磷含量的变化相对较小,埋深为0~10 cm、10~20 cm、20~30 cm和30~50 cm土壤中全磷含量随土壤藻结皮发育年龄的增加,分别依次增加了4.7%、10.9%和20.3%,3.1%、7.8%和18.8%,4.8%、6.3%和17.5%,1.6%、4.9%和13.1%。
上述试验结果表明,在埋深为0~50 cm土壤中总磷含量变化较小的情况下,其中土壤中速效磷含量所占比例随土壤藻结皮发育年龄的增加而增加,表明土壤中有更多磷素能被植物利用而增加了植物的生物量,从而加快了退化草地的修复。
2.5 土壤藻改良过程中退化草地植物群落组成的变化
接种土壤藻对退化草地中植物群落组成和各物种重要值的影响,见表2。
由表2可知:在接种土壤藻之前,退化草地植物群落组成中草本植物主要有本氏针茅、沙打旺、碱蓬、芨芨草、披碱草、羊草,小灌木有油蒿;
接种土壤藻改良3年后,退化草地中增加了草本植物沙蓬、灌木羊柴、沙柳和柠条,而未接种的退化草地没有后续入侵生物出现;
在土壤藻改良3年的过程中,退化草地中这些植物物种的重要值发生了很大变化,本氏针茅、沙打旺、碱蓬、芨芨草、披碱草、羊草和油蒿的重要值分别由接种前的0.241、0.219、0.154、0.101、0.097、0.045和0.143,变为接种后的0.194、0.087、0.109、0.067、0.196、0.129和0.055;
在改良的第三年,退化草地中增加的4种植物物种沙蓬、羊柴、沙柳和柠条的重要值分别为0.103、0.034、0.021和0.005。

表2 土壤藻结皮对退化草地中植物群落组成和各物种重要值的影响
上述试验结果表明,接种土壤藻明显改变了退化草地植物群落物种的重要值。
2.6 土壤藻改良对退化草地中植物群落特征值的影响
不同土壤藻改良年龄内退化草地中植物群落特征值的变化,见表3。
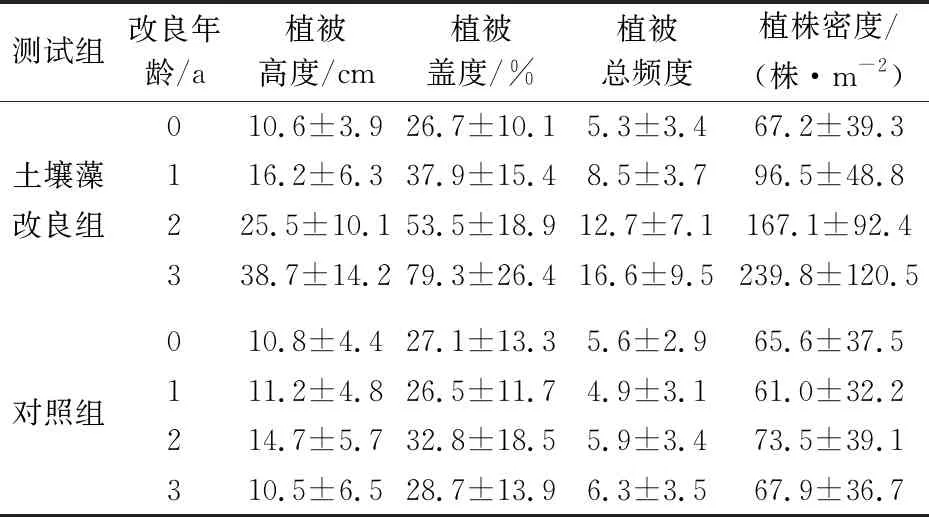
表3 不同土壤藻改良年龄内退化草地中植物群落特征值的变化
由表3可知:在土壤藻改良的3年内,退化草地中植被高度由接种前的10.6 cm增加至38.7 cm,植被盖度由26.7%增加至79.3%,植被总频度由5.3增加至16.6,植株密度也由67.2 株/m2增加至239.8 株/m2;
植被高度、植被盖度、植被总频度和植株密度的增幅分别为265%、197%、213%和257%,其中植被高度的增幅最大,植株密度次之,即使增幅最小的植被盖度,增加幅度也达到197%;未接种土壤藻的对照组退化草地中植被高度、植被盖度、植被总频度和植株密度的变化较小,只在土壤藻改良的第二年这些植物群落特征值有一个小幅度增加,这可能是因为当年降雨量增加,植被生长相对较好所致。
2.7 土壤藻改良过程中退化草地植物群落物种多样性的变化
不同土壤藻改良年龄内退化草地中植物群落物种多样性指数的变化,见表4。

表4 不同土壤藻改良年龄内退化草地中植物群落物种多样性指数的变化
由表4可知:接种土壤藻后,随着土壤藻结皮发育年龄的增加,退化草地中植物群落物种的多样性指数、丰富度指数和均匀度指数均随之升高,且增加幅度较大;
土壤藻改良3年后,退化草地中植物群落物种的多样性指数、丰富度指数和均匀度指数分别由1.412、0.488和0.435增加至2.358、0.890和0.687,增幅分别为67.0%、82.4%和57.9%;
而未接种土壤藻的对照组退化草地中,植物群落物种这三个指数在初始值的附近变化。
上述试验结果表明,退化草地接种土壤藻后,植物群落物种的多样性更为丰富,分布更加均匀。
2. 8 土壤藻改良对退化草地中植物产草量的影响
不同土壤藻改良年龄内退化草地中植物生产量的变化,见表5。

表5 不同土壤藻改良年龄内退化草地中植物生产量的变化
由表5可知:退化草地中植物鲜重和干重随土壤藻改良年龄的增加而增加;经过一年的土壤藻改良,每平方米退化草地中增加植物鲜重80.3 g和植物干重27.6 g;
土壤藻改良的第二年增加也较快,每平方米退化草地中植物鲜重和干重的增加量达到188.4 g和51.8 g;
改良的第三年每平方米退化草地中植物鲜重和干重的增加量最高,分别为植物鲜重253.5 g和植物干重103.7 g;
而未接种土壤藻的退化草地,其生物量变化较小,只在第二年有一定的增加,比第一年增加植物鲜重29.6 g和植物干重10.3 g。此外,表5也显示了不同土壤藻改良年龄内退化草地中植物的干鲜比,尽管接种土壤藻的退化草地中植物鲜重一直处于增加状态,但是植物干鲜比却呈现先增加后减少的变化趋势;
而未接种土壤藻的对照组退化草地,植物的干鲜比除了改良的第三年下降外,前两年均增加,只是增加幅度较小。
3. 1 土壤藻改良对退化草地土壤肥力的影响
土壤含水量是影响土壤肥力的一个重要因素。土壤水分不仅影响土壤的理化性质,制约土壤中矿质营养的溶解、迁移和转化,而且还影响着土壤微生物的活动,制约土壤肥力的变化[21]。土壤含水率的变化是因为随着土壤藻结皮的发育,草本植物物种多样性和生物量增加,地表植被盖度加大,降低了土壤水分的蒸发;
同时,植物根系腐烂后,增加了土壤的孔隙度,为水分的渗透留下了通道[22];
另外,由于土壤藻形成的生物结皮具有很强的保水能力,能将短时间降雨和夜晚的露水吸收、下渗而保存在土壤中[17,23]。
土壤有机质含量是土壤肥力的重要指标。土壤藻改良退化草地的土壤中有机质含量增加,主要是因为随着土壤藻的生长发育,代谢活动加强,蓝藻通过光合作用将环境中的CO2转化为碳水化合物,同时固氮蓝藻通过固氮作用将大气中植物不能直接利用的N2转化为化合态氮,并通过生物的代谢作用合成含氮有机化合物[15]。土壤藻类通过光合作用、固氮作用和其他代谢作用改善土壤环境,为植物的生长创造了有利条件。草本植物在退化环境中得以继续生长发育,随着植物地上生物量的增加,光合作用强度加大,同时也增加了有机质向土壤中的输入量,使得土壤中有机质含量急剧增加。另外,植物根系和藻类生物结皮主要集中分布在土壤表层,因此植物根系、草本植物地上残体和土壤藻死亡后的藻体对土壤有机质的贡献主要作用在土壤表层。土壤中有机质含量随土壤埋深的增加而递减,可能是因为随着土壤深度的增加,植物地上残体、死亡藻体和根系数量逐渐减少,从而导致土壤中有机质含量的下降。土壤团聚体是表征肥沃土壤的指标之一,而土壤有机质有利于土壤团聚体的形成,并可增加土壤的孔隙度[22]。另外,土壤肥力越强,土壤团聚体越多,而团聚体越发达的土壤保水保肥能力就越强,也越有利于退化草地的生态修复[24]。
如前文所述,固氮蓝藻能将大气中N2转化为能被植物吸收利用的化合态氮而固定下来。因此,随着土壤藻结皮的发育和植物的生长,土壤中氮素含量逐渐增加,而土壤中全氮含量的增加说明退化草地正在逐渐恢复。此外,有研究还发现,土壤脲酶活性随着土壤藻形成的生物结皮的发育而增强[25],而土壤脲酶可以将土壤中含氮有机化合物水解为氨氮化合物供植物吸收利用。因此,土壤中速效氮含量也随之增加。
磷与氮一样,也是植物生长发育必需的营养元素,是土壤肥力的重要指标。土壤磷酸酶在土壤磷素转化和循环中起着重要作用[26],而土壤磷素转化效率与土壤磷酸酶活性成正相关关系。因此,可以利用土壤磷酸酶的活性来评价土壤磷素转化的强弱。前期的研究也证明,土壤磷酸酶活性随着土壤藻结皮的发育而增强[25],这表明土壤藻的生长和发育提高了土壤磷酸酶的活性,而土壤磷酸酶可以加快催化土壤中磷素的转化,使土壤中速效磷含量增加。
实际上,在土壤中微生物是参与碳、氮、磷、硫等元素转化的主要驱动力,对土壤植物生态系统中的物质循环和能量转化起着决定作用[27]。因此,随着土壤藻生物量的增加,土壤藻不仅加快了土壤中磷素的转化,也加快了磷的循环。这与本研究结果中退化草地土壤中速效磷含量随土壤藻结皮的发育而增加相一致。退化草地土壤中全磷的含量变化不大,这可能是因为磷在土壤中较为稳定,其含量尽管与土壤有机磷的净矿化作用、土壤磷素的微生物和非微生物固定有关,但是其含量主要取决于土壤母质的类型和质地[28]。
3.2 土壤藻改良对退化草地中植物群落物种多样性的影响
土壤藻改良3年后,随着退化草地逐渐恢复,植物群落的组成也发生了相应变化。在接种土壤藻之前,退化草地中主要以本氏针茅、沙打旺、碱蓬和芨芨草等草本植物为主,但也分布有小灌木油蒿;
随着土壤藻结皮的发育,土壤营养和生态小环境得到改善,之前长势较差的草本植物生长加快,同时其他植物逐渐入侵,导致原来的几种优势植物在群落中的比重下降。特别是之前生长很好的油蒿,在土壤藻改良3年后,其在植物群落中的地位明显下降,几乎要从研究区消失,这可能与油蒿的生长特性和生态位有关。油蒿主要生长在其他物种较少的干旱和半干旱地区,常常作为沙地建群种在流动沙丘上旺盛生长,但随着流动沙丘被固定,其生长能力也随之下降,最后死亡而被其他生长旺盛的植物所替代。同样,随着土壤藻结皮的发育,退化草地中改良前的主要优势植物沙打旺和芨芨草的地位逐渐下降,在土壤藻改良3年后,退化草地中植物群落由本氏针茅-沙打旺群落演替为披碱草-本氏针茅群落,后续入侵植物沙蓬经过3年的快速生长和繁殖,在植物群落中占有了很重要的地位。
土壤藻结皮的发育不仅改变了植物群落的植被组成及其重要值,而且也影响着植被的高度、盖度、总频度和植株密度等主要特征值,土壤藻改良区植被的高度、盖度、总频度和密度明显高于对照区植被的相应值,且随着土壤藻结皮的发育,这些植物群落的特征值也逐渐增加。
作为草地生产力的主要数量指标,植被的高度和盖度决定着草地的产草量,而植被频度和植株密度在某种程度上也影响着草地的生产力。植被频度是植物物种丰富度及其空间分布均匀程度的一项综合指标,植被频度越高,说明植物物种的丰富度及其空间分布均匀度越高[29-30]。本研究的结果表明,退化草地植物的丰富度和均匀度随土壤藻结皮的发育而升高。从土壤藻结皮发育影响植物群落特征值的结果可知,土壤藻结皮对植被高度和植株密度的影响大于对植被盖度和植被频度的影响,也就是说,土壤藻结皮在垂直方向上促进了植物向更高空间生长,在水平方向上增加了单位面积的植株数量。植物群落中特征值的变化表明,土壤藻改良后退化草地植物发育越来越好。由此可见,土壤藻结皮的生长明显促进了退化草地植被的发育,加速了退化草地的生态恢复。
植物群落物种的多样性指数直接反映植物群落的结构特征。未接种土壤藻的对照组退化草地,植物群落物种多样性水平较低,植物群落结构简单,物种组成少,这可能是因为草地生境条件日益恶化,植物群落物种组成减少且分布不均;
接种土壤藻的退化草地,其植物群落物种多样性指数快速升高,表明植物群落结构越来越复杂,物种组成越来越丰富。如前文所述,土壤藻结皮的发育提高了土壤含水率,增强了土壤酶的活性,增加了土壤肥力,改善了退化草地的生境;
而得到改善的生态环境不仅影响着土壤藻的生长发育和繁殖,同时也影响和改变着退化草地植物的形态结构和生理生化特性,促进它们更加适应变化了的环境条件而快速生长和发育,并在退化草地均匀分布;
当退化草地环境得到进一步改善时,随风力飘落或者被动物带入的植物种子或营养枝在退化草地生根发芽,使得退化草地植物物种增多,植物群落物种丰富度指数增加,退化草地植物群落的物种更加丰富。
接种土壤藻后,退化草地生物量明显增加的原因可能是因为土壤藻结皮提高了土壤酶活性,提升了土壤活力,增加了土壤含水率和土壤肥力,从而改善了退化草地植物生长的环境条件,促进了退化草地植物的生长,进而导致退化草地植物年产草量的快速增加。群落植物干鲜比反映了植物含水量,对土壤藻改良过程中退化草地植物干鲜比的分析,说明群落植物含水量发生了相应的变化。有研究表明,群落植物干鲜比值受演替不同阶段物种组成差异和降雨量变化等方面的影响,其中物种组成差异起决定作用,禾本科干鲜比最高,其次是菊科和豆科[31]。前文研究结果也表明,随着土壤藻结皮的发育,退化草地中植物群落物种的丰富度增加,有一些小灌木物种入侵,但是相对于草本植物来说,小灌木的相对生物量增加较小,导致植物干鲜比变化不明显。另外,尽管在土壤藻改良的第二年研究区有一定的降雨,但是改良草地和对照草地植物的干鲜比没有出现相同趋势的变化,前者有轻微降低,而后者却出现轻微升高,说明在改良草地植物体中水分持有量比对照草地植物高。
在退化草地接种土壤藻,培植土壤藻结皮,随着土壤藻结皮的发育,土壤的水分、有机质、氮和磷含量都呈现增加趋势,这些环境因子的变化将有利于退化草地植物的生长,使得植物生物量快速增加。退化草地植物生长微环境得到了改善,为后续入侵的沙蓬,羊柴、沙柳和柠条等草本植物的生长创造了有利条件,从而改变了退化草地的植物群落组成。同时,土壤藻改良区植被高度、植被盖度、植被总频度和植株密度等主要特征值明显增加,退化草地中植物群落物种的多样性指数、丰富度指数和均匀度指数也大幅提升,且退化草地中植物的鲜重和干重随土壤藻结皮的发育而增加。这些参数的变化都表明土壤藻结皮可以改良退化草地,加速退化草地的生态系统恢复。
猜你喜欢结皮速效草地NaCl与Na2SO4风沙土盐结皮力学特性比较分析∗新疆大学学报(自然科学版)(中英文)(2022年3期)2022-06-04黄土高原不同发育阶段生物结皮的导水和持水特征草业学报(2021年6期)2021-06-28草地上的事幼儿100(2020年31期)2020-11-18黄土高原典型生物结皮对坡面产流产沙过程的影响水土保持通报(2020年6期)2020-03-10Laughing song疯狂英语·初中版(2019年4期)2019-09-10草地小太阳画报(2018年6期)2018-05-14土壤风蚀环境下盐结皮研究及展望∗新疆大学学报(自然科学版)(中英文)(2018年4期)2018-02-10不同杀螨剂对柑桔红蜘蛛速效与持效性防治田间试验现代园艺(2017年21期)2018-01-03土壤速效钾测定方法的影响因素绿色科技(2016年16期)2016-10-11施用不同有机物料对山地果园红壤速效氮磷钾含量的影响亚热带资源与环境学报(2015年1期)2015-01-22