王凯悦 张仲富 王好才 展鹏飞 宋维峰 陆 梅 王 行
(1. 西南林业大学云南省高原湿地保护修复与生态服务重点实验室,云南 昆明 650233;
2. 西北大学生命科学学院,陕西 西安710069;
3. 西南林业大学国家高原湿地研究中心/湿地学院,云南 昆明 650233;
4. 福建师范大学地理科学学院,福建 福州 350007;
5. 西南林业大学生态与环境学院,云南 昆明 650233)
微生物是土壤物质形成与转化的关键驱动力[1-2],同时也是地球元素循环的引擎[3],对整个生态系统的稳定具有重要意义。研究土壤微生物群落空间分布格局以及功能特征有助于进一步阐明土壤生态功能[4]。微生物群落结构及功能变化势必导致土壤功能的强弱、指向性趋势发生变化。由于土壤微生物对环境改变十分敏感,因此可以通过对微生物群落特征变化的研究得知土壤状况的改变[5]。当前,随着人口数量不断增大,土地利用类型发生了极大的变化,人们对土地利用的关注度持续上升,原生土地被人为干扰程度相较于其他土地方式的转变更加明显,其中农田耕作是自然土壤最大的干扰源[6]。大量研究表明,人为因素的干扰改变土壤微生物群落结构特征[7]。如靳振江[8]以太湖地区典型的潴育型水稻土-黄泥土为试验田,将采样区分为不施肥小区、单施化肥小区、化肥配施秸秆小区和化肥配施猪粪小区,对各小区进行对比分析后发现,农田单施化肥导致土壤微生物多样性明显减少;
祖智波等[9]也在其研究中指出,机械耕作土壤中的好气性细菌、厌气性细菌、放线菌数量都低于少耕土壤。当前,绝大多数研究以现代机械耕作下的水稻土作为研究对象,且重点关注不同施肥方式的影响,传统农耕下的土壤微生物群落结构及潜在功能变化特征尚不明确。
哈尼梯田是我国西南山地农业耕作方式的典型代表,广泛分布于云南省红河州境内,州内水稻耕作面积约有12 667 hm2[10]。哈尼梯田水稻种植区是哈尼族人由森林开垦而来,他们创造性地利用了水热条件适宜的中山缓坡地带,建造并维持了以水梯田为主体、具有独特生态功能的亚热带山地农业景观[11]。近些年对哈尼梯田土壤的研究大多集中于土壤重金属[10]、土壤养分[12-13]等方面。对于揭示传统耕作方式下土壤微生物的特征及其与附近未受干扰原生土壤——森林土作对比的研究还未见报道。因此,本研究以哈尼梯田水稻土与森林土作为研究对象,采用高通量技术对比研究了2种土壤中的微生物群落结构及潜在功能特征,旨在揭示传统人为耕作方式对哈尼梯田水稻土壤微生物群落结构及功能变化的影响,以期为进一步了解人为干扰下的土壤生态功能演变提供科学依据和理论支持。
哈尼梯田核心区位于云南省元阳县(102°27′~103°13 ′E,22°49′~23°19 ′N),面积2 189.88 km2,最高海拔3 074.3 m,最低海拔105 m[14],属哀牢山南部,为中山深切割地貌类型,北部与元江水系相邻,南部受藤条江的切割[15],地形由西北向东南倾斜[16]。境内地势陡峻,坡度在25°以上的土地占土地总面积的59.1%。研究区属于山地季风气候,雨热同季、冬无严寒、夏无酷暑,立体气候显著[17],年平均温度11.6 ℃,年平均降水约3.2 mm/d。以森林、村庄、梯田、水系为物质载体,梯田的开垦及种植历史有1 300多年[11-19]。
2.1 样地设置与样品采集
采样区域位于红河哈尼族彝族自治州的东部元阳县西南部,采用大规模采样调查,采样区面积约226.56 km2,沿海拔梯度进行采样,样品主要分布于元阳梯田景区内、绿春县、甲寅镇附近。水稻土采集完整梯田内的土壤,森林土采集水稻土附近森林内未经人为耕作、自然状态下的土壤。具体地,在每一处水稻土采样点附近相应设置具有典型森林特征的一处森林土采样点,沿海拔梯度共采集10处水稻土,分别对应周围10处森林土采样点,保证各采样点之间的配对关系,使样品之间具有可比性。采样区森林中多生长杉树纯林、云南松纯林、杉树-云南松混交林这类亚热带常绿阔叶林。水稻土与森林土样品采集时间为水稻生长盛期(2018年6月份)。在水稻土采样区内沿海拔梯度设置10个5 m×5 m的样方,选取稻田中心土壤进行取样(避免边缘效应),每个样方均利用对角线多点(共选取5点)混合取样法采集0~15 cm表层土壤[19]。森林土的采样点选取距离水稻土采样点较近的森林中心区域的林下土壤,采样方式同水稻土。结合现场分析研究以及高分卫星遥感影像数据历史判读,确保所选森林土没有经过人为耕作管理,通过与水稻土进行对比研究,能够真实反映人为耕作对原生土壤微生物群落结构的干扰响应特征。研究区域的气候数据,包括每个采样点的年平均降水量和年平均温度等均来自WorldClim全球气候数据集[20]。
2.2 土壤样品理化指标分析
土壤理化性质的测定主要参考鲍士旦的《土壤农化分析》[21]。土壤总氮(TN)采用半微量凯氏定氮法测定;
总碳(TC)采用重铬酸钾氧化法测定;
总磷(TP)采用消解钼锑抗分光光度法测定;
土壤中有效磷(Olsen-P)采用碳酸氢钠浸提钼锑抗比色法测定;
土壤pH采用酸度计法测定;
利用环刀中的土样可一次性测定土壤容重等基本理化指标;
土壤导电率(Conductivity)采用电极法进行测定[5];
淹水深度(FD)使用米尺测量。
2.3 土壤微生物DNA提取和16S rRNA扩增子测序
土壤DNA提取使用CTAB法,共提取20份DNA,其中包括水稻土10份,森林土10份。DNA的质量用Nanodrop 2000c检测,后利用引物515F(5′-GTGCCAGCMGCCGCGG-3′)和 907R(5′-CCG-TCAATTCMTTTRAGTTT-3′)对细菌16S rRNA进行PCR扩增[22]。反应体系为:5 U/μL Taq酶 0.25 μL,10×PCR反应缓冲液5 μL,2.5 mmol/L dNTP 4 μL,10 μmol/L正反向引物各1 μL,DNA模板1 μL(约20~50 ng),加水至50 μL。扩增条件为:
95 ℃预变性5 min;
然后在95 ℃ 30 s,55 ℃ 30 s,72 ℃ 60 s,共进行24个循环;
最后在72 ℃延伸8 min。获得扩增产物后,利用Agarose Gel DNA Fragment Recovery Kit试 剂 盒(Ta KaRa)切胶纯化,并溶于100 μL Dnase-free水中。PCR产物的纯度和浓度使用QuantiFluor TM-ST蓝色荧光定量系统(Promega,美国)进行检测定量。在文库质检后,使用Illumina MiSeq测序仪进行测序[23]。
2.4 数据处理与统计分析
采用QIIME(Version 1.7.0)对细菌16S rRNA进行原始数据的处理[24];
应用软件R语言的vegan包计算样品Alpha多样性,使用ggplot2包对将数据进行可视化,使用AI(Adobe illustrator)图片处理工具对所做图形进行美化,使用SPSS 22.0进行数据差异性检验。Alpha多样性包括种群丰富度、Chao1指数、Simpson指数、Shannon指数、ACE指数;
对样品Beta多样性的分析在多元统计分析软件Canoco 5.0中进行,并通过Weighted Unifrac PCoA分析得出微生物群落结构在样品间的差异。将代表性微生物序列对比RDP参考数据库可以得到物种信息,对物种门、属水平进行分析后获得样本优势物种相对丰度信息,随后通过Venn图得出样本中的核心群落并对核心群落进行功能分析比较。在OmicShare(http://www.omicshare.com/tools)平台上进行典型相关分析(CCA),得出对环境变化敏感的微生物,由2组样本的Circos图获得核心微生物群落的丰度;
在Galaxy在线分析平台进行LEfSe差异统计分析,找到组间生物标志物。FAPROTAX特别适用于对环境样本的生物地球化学循环过程(特别是碳、氢、氮、磷、硫等)进行功能注释预测,因此本研究采用FAPROTAX v1.1进行微生物功能预测分析。利用博大研究所开发的在线工 具 Morpheus(https://software.broadinstitute.org/morpheus/)绘制微生物功能特征的热图(Heatmap),由图进一步判断微生物的功能差异。
3.1 土壤理化性质分析
由表1可知,水稻土与森林土相比,土壤中的总碳、总磷明显下降,分别下降了59.15%、36.59%,表明由于反复耕作使得土壤有机质含量下降[25-26];
人为耕作对土地的改造(如施加磷肥及农田灌溉等),使得土壤有效磷含量和淹水深度明显增加;
水稻土与森林土pH差异不显著,均呈酸性,且电导率也无明显差异;
水稻土中总氮含量略有下降但差异不显著。
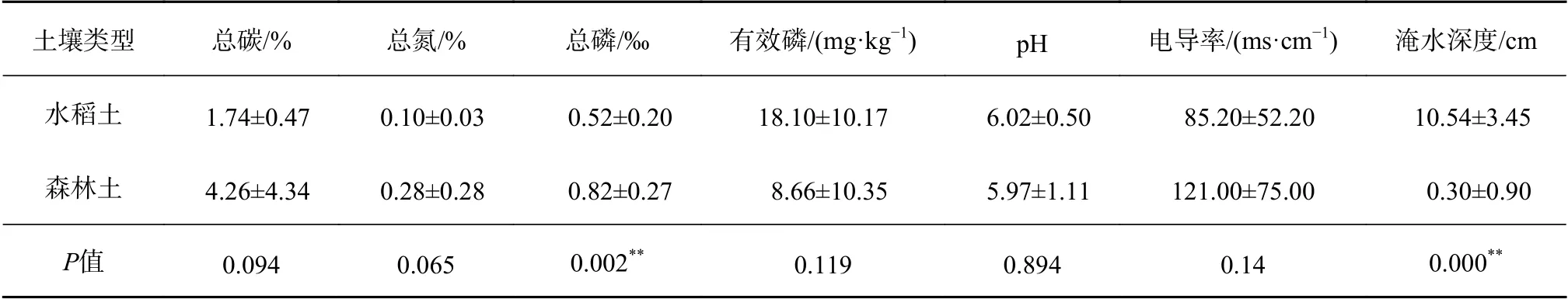
表 1 水稻土与森林土壤理化性质特征Table 1 The physical and chemical properties of paddy soil and forest soil
3.2 土壤微生物群落多样性分析
由表2可知,森林土微生物的物种丰富度为17 494个,人为耕作影响下的哈尼梯田水稻土微生物的物种丰富度增加到23 023个。与森林土相比,水稻土Chao1指数、ACE指数显著分别增加了107.32%和117.96%;
水稻土壤微生物的Shannon指数较森林土有小幅增加,而Simpson指数在两者之间没有显著差异。
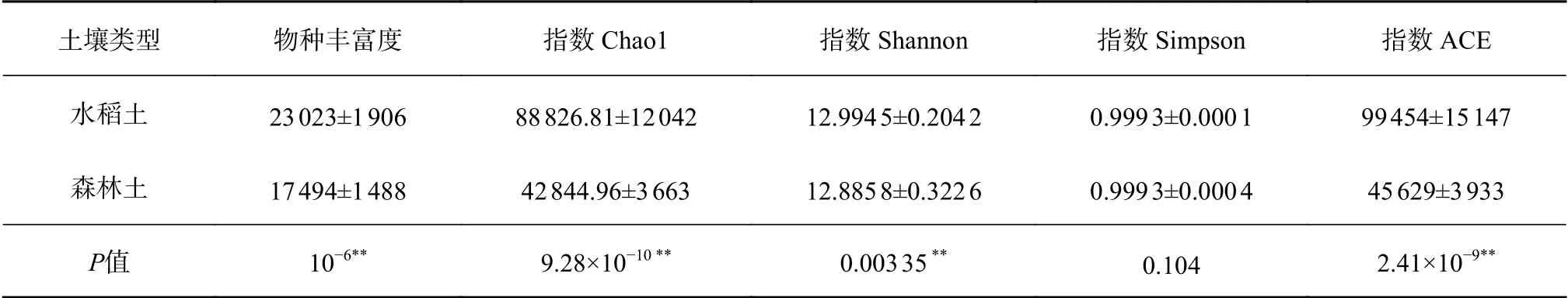
表 2 水稻土与森林土微生物Alpha多样性指数Table 2 Alpha diversity index of microorganisms in paddy soil and forest soil
样品的取样深度可以通过稀释曲线来评价测序量能否覆盖所有类群,其结果见图1。随着测序深度的增加,土壤样品稀释曲线呈上升趋势,说明样品的物种多样性较高,但仍有较多未被检测到的物种,水稻土的物种丰度明显大于森林土。同时,图1b的Rank-Abundance曲线表明,森林土曲线相比水稻土更加平缓,因此森林土中的微生物物种分布更加均匀。
微生物群落的Beta多样性采用Weighted Unifrac PCoA进行分析,结果见图2。由图2可知,虚线圈分别涵盖了大部分的采样点并聚类明显、区分度较高,表明水稻土与森林土微生物群落组成存在较大差异。其轴1和轴2的解释度分别为44.32%和5.25%,累计贡献率达到49.57%。森林土位于轴1的正端,水稻土集中分布于轴1负端,表明不同土壤样品之间微生物群落结构的差异主要由排序轴1决定。
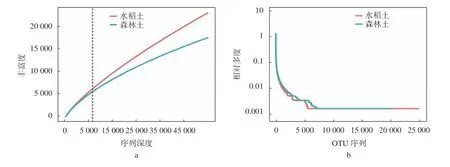
图 1 物种相对丰度稀释曲线Fig. 1 Relative abundance dilution curve of species

图 2 水稻土与森林土壤微生物群落结构Beta多样性分析Fig. 2 The Beta diversity of microbial community structure in paddy soil and forest soil
3.3 土壤微生物群落结构分析
3.3.1微生物分布特征分析
Venn图可表示样品中微生物数目组成的相似性以及重叠情况,水稻土样本中共检测到959个微生物OTU单元,占55.08%,森林土样本含有1 186个微生物微生物OTU单元,占74.40%(图3a、3b),人为耕作活动对土壤的干扰导致土壤核心微生物群落数量减少了19.14%。进一步将水稻土与森林土的核心微生物群落进行分析,得出其重叠状况(图3c),两者重叠的OTU有106个,其优势物种为厌氧粘细菌(Anaeromyxbacter)以及黄单胞杆菌(Xanthomonadales),重叠率仅占总数的5.20%。水稻土特有OTU单元853个,包括拟杆菌(Bacteroidetes)、硝化螺旋菌(Nitrospirae)等,森林土特有OTU单元1 080个,以浮藻菌(Planctomycetes)、β-变形菌(Betaproteobacteria)等优势微生物为主。

图 3 水稻土与森林土壤微生物OTU分布Venn图Fig. 3 Venn diagram of paddy soil and forest soil microbial OTU distribution
3.3.2门水平群落结构分析
以门为分类标准对样品进行分析(图4a),森林土以放线菌门(Actinobacteria)为优势菌门(28.94%),其他优势菌门包括绿弯菌门(Chloroflexi,21.16%)、浮藻菌门(20.43%)和硝化螺旋菌门(7.98%)。水稻土以绿弯菌门为优势菌门(37.64%),其他优势菌门包括拟杆菌门(11.54%)、硝化螺旋菌门(8.09%)和放线菌门(8.03%)等。与森林土相比,水稻土中的放线菌门和浮藻菌门显著减少,分别减少了20.92%和14.53%;
而绿弯菌门和拟杆菌门显著增加,分别增加了16.48%和6.28%。
3.3.3属水平群落结构分析
以属为分类标准对样品进行分析(图4b),水稻土以地杆菌(Geobacter)为优势菌属(21.01%),其他优势菌属包括螺旋体属(Spirochaeta,18.00%)、慢生根瘤菌属(Bradyrhizobium,17.23%)、厌氧粘细菌(12.89%)以及同营养杆菌(Syntrophorhabdus,10.27%)。森林土中以慢生根瘤菌属为优势菌属(26.66%),其他优势菌属有三角变形杆菌(Deltaproteobacterium,18.87%)、嗜酸栖热菌属(Acidothermus,13.56%)、Haliangium(11.39%)和布氏杆菌属(Bryobacter,11.38%)。在属水平进行对比发现,水稻土样品中的厌氧粘细菌和慢生根瘤菌属显著增加,分别增加了16.72%和15.12%;
而嗜酸栖热菌属显著减少,减少了12.80%。
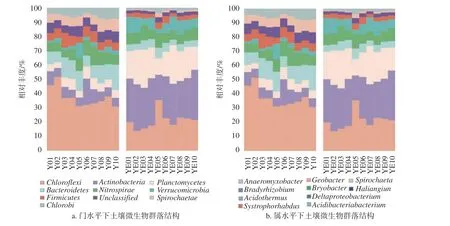
图 4 土壤微生物在门和属分类水平上的群落结构Fig. 4 Soil microbial communities at the level of phylum and genus
3.3.4关键指示微生物分析及相关功能列表
对哈尼梯田水稻土与森林土的核心微生物进行分析,揭示关键指示微生物的物种信息。以属为分类标准得到在水稻土中的优势菌为地杆菌属(27.85%)、螺旋体菌属(20.67%)和厌氧粘细菌(20.13%);
在森林土中的优势菌为慢生根瘤菌属(29.45%)和嗜酸栖热菌属(19.70%)(图5)。菌群功能详见表3。

表 3 核心微生物相关功能Table 3 Core microbial and their related functions

图 5 水稻土与森林土属水平核心微生物群落分布特征Fig. 5 Distribution characteristics of core microbial communities of order horizontal in paddy soil and forest soil
3.3.5组间LDA差异贡献分析
为进一步验证传统农耕方式下的哈尼梯田水稻土与森林土微生物组成的相似性和差异性,采用LEfSe进行差异统计分析,找到组间具有统计学差异的生物标记物(Biomarker)。设定LDA score=4.1时进行LDA非参数秩和检验,通过检验找出哈尼梯田水稻土与森林土样品中微生物在不同分类水平上可能存在的生物标志物。由图可以看出,森林土有12个生物标志物,水稻土有8个生物标志物,地杆菌是水稻土中影响最大的生物标志物(图6),慢生根瘤菌是森林土中影响最大的微生物标志物。该研究结果与图5得到的结果具有一致性。
3.4 土壤微生物群落结构与环境因子的相关性分析
微生物群落结构与环境因子的典型相关分析(CCA),结果见图7。水稻土轴1的解释度为54.47%,轴2的解释度为15.77%,2个轴累计贡献率达到70.24%。淹水深度对水稻土壤微生物的分布影响最为显著,总磷与土壤微生物群落的相关性最高。在属分类水平上,地杆菌(Geobacter)和螺旋体属(Spirochaeta)等分布比较集中,且与海拔、总磷等环境因子所成夹角较小,表明土壤中的这些微生物对土壤环境因子的响应较为敏感。森林土轴1的解释度为34.06%,轴2的解释度为30.95%,2个轴累计贡献率达到65.01%。总磷和海拔对森林土壤微生物分布影响较为显著,淹水深度和pH与土壤微生物群落相关性最高。以属为分类水平,慢生根瘤菌(Bradyrhizobium)、布氏杆菌属(Bryobacter)和嗜酸栖热菌属(Acidothermus)等分布比较集中且与淹水深度、海拔等环境因子所成夹角较小,表明土壤中的这些微生物对土壤环境因子的响应较为敏感。

图 6 水稻土与森林土组间线性判别分析Fig. 6 Linear discriminant analysis between paddy soil and forest soil
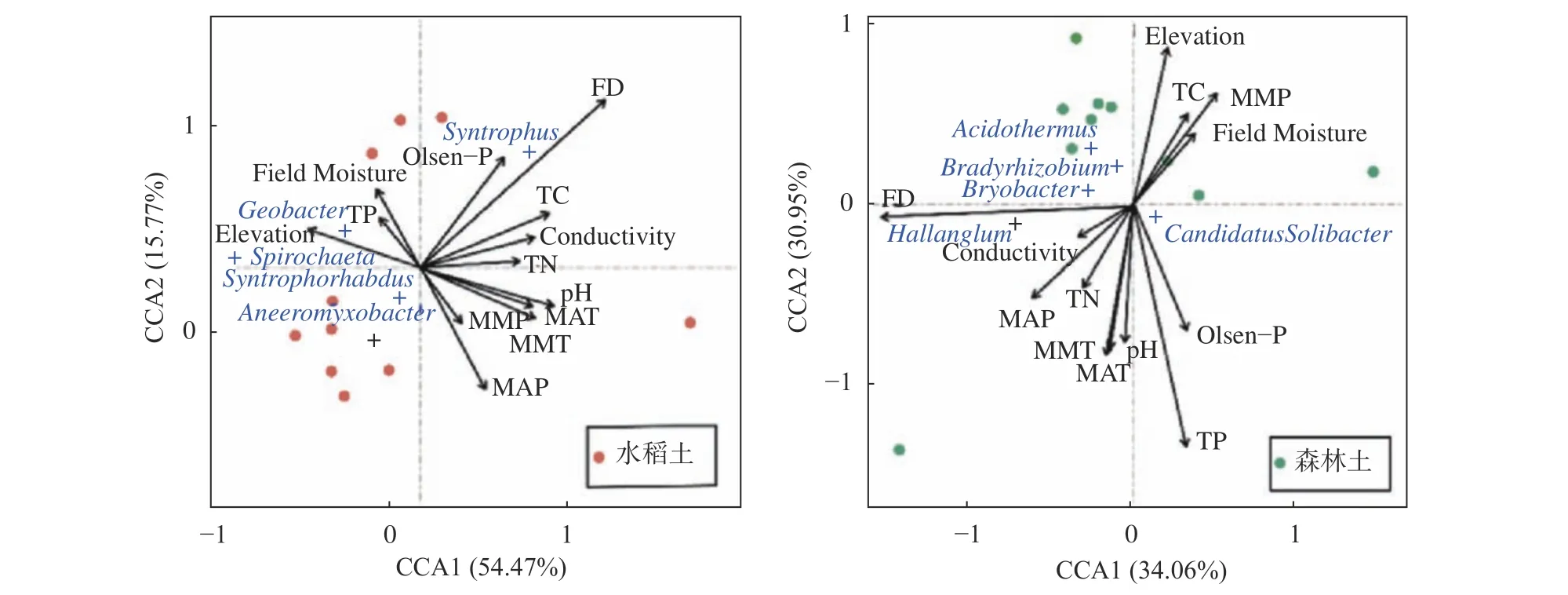
图 7 微生物属水平群落结构与环境因子的CCA分析Fig. 7 Canonical correspondence analysis of microbial community structure and environmental factors at the genus level
3.5 土壤微生物功能特征分析
利用Heatmap图展示土壤微生物功能特征(图8)。由图8可知,2样品聚类明显,水稻土样品中所含菌群的功能主要涉及铁呼吸作用、硫化物呼吸作用、好氧亚硝酸盐氧化、发酵作用等与元素循环有关的功能潜能;
而森林土样品中菌群的功能主要涉及纤维素降解、芳香烃降解、木质素分解等与有机质降解有关的功能潜能。

图 8 微生物功能特征分析热图Fig. 8 The heatmap for analysis of microbial functional characteristics
4.1 讨论
4.1.1哈尼梯田传统农耕作用下水稻土与森林土微生物群落多样性变化
土地利用方式的不同显著影响土壤微生物群落多样性。张杰等[37]对鄱阳湖湿地不同土地利用方式下土壤微生物群落功能多样性进行研究发现,人为耕作使土壤微生物群落多样性显著下降,通过“退田还湖”等措施可恢复土壤微生物的多样性。秦红等[38]对三峡库区消落带土壤细菌和真菌多样性进行研究得出结论,与弃耕地相比,人工耕地微生物多样性受到破坏,提倡进行人工生态修复以恢复三峡库区的土壤微生物多样性。前人的研究结果普遍揭示人为耕作影响使得土壤微生物的多样性低于未被人为干扰的土壤。然而在本研究中发现,哈尼梯田的水稻土的Alpha多样性显著高于森林土,表明在传统农耕方式下水稻土壤中的微生物数量相较于未经人为耕作的森林土多样性显著增加,传统耕作中有机肥与化肥配施很可能是造成微生物多样性增加的主要因素。已有研究表明,耕作时使用有机肥与化肥配施的模式有利于提高土壤微生物多样性[39],与当前机械耕作(仅施加化肥)对土壤微生物多样性造成胁迫形成对比,添加有机肥料养分有利于改变土壤微生物群落碳源代谢模式,提高微生物群落碳源利用能力,从而导致土壤微生物多样性提高。徐万里等[39]提出Shannon指数与纤维素分解菌之间显著正相关性,土壤中微生物种类丰富度与纤维素分解菌密切相关。哈尼梯田水稻土的Shannon等多样性指数较森林土均有所增加,可推测水稻土的微生物群落多样性高于森林土。化肥与有机肥长期配施可提高水稳性团聚体的稳定性及各粒级团聚体内有机碳含量[40];
袁红朝等[41]提出氮磷钾配施和化肥配施可显著提高土壤细菌多样性,而后采用T-RFLP技术对同地带的水稻土不同施肥下的微生物群落变化进行研究并证实这一想法。前人及我们的研究表明,传统耕作方式下的水稻土壤,通过采用化肥有机肥配施模式,可显著维持甚至提高土壤微生物的多样性。
4.1.2哈尼梯田传统农耕下形成的水稻土壤微生物群落结构变化
通过对哈尼梯田水稻土与森林土核心菌群研究发现,传统耕作方式下的水稻土壤微生物的核心菌群为地杆菌、螺旋体菌和厌氧粘细菌,作为对照的森林土微生物核心菌群为慢生根瘤菌和嗜酸栖热菌,表明化肥-有机肥配施方式下的水稻土核心微生物多为一些酸杆菌属、螺旋菌属。通过CCA可以看出,pH和总磷与土壤微生物群落的相关性最高。由于水稻土壤淹水深度显著高于森林土,所形成的淹水环境使得地杆菌的丰度大大增加。Lovley等[42]在其研究中指出,在各种厌氧环境中的主要微生物种群为地杆菌科微生物;
朱超[43]研究了淹水环境对水稻土壤微生物群落结构的影响,得出厌氧环境导致水稻土壤中地杆菌与厌氧粘细菌数量显著增加的结论。
进一步对水稻土中的微生物群落进行探究,发现地杆菌和厌氧粘细菌都是重要的异化Fe(Ⅲ)还原菌,具有降解土壤芳香族有机物、重金属污染修复等功能潜能。研究区所处西南地区,有大量有色金属成矿带[10],土壤重金属背景值偏高可能是土壤中地杆菌与厌氧粘细菌数量偏高的原因之一;
传统耕作方式下的有机肥与化肥配施对土壤也会产生一定污染[44],如土壤中有效磷等含量增加,地杆菌、厌氧粘细菌的存在有利于降低土壤污染程度。人为耕作方式下的水稻土壤中地杆菌和厌氧粘细菌的存在对土壤中重金属污染修复、土壤生态系统稳定以及农业生态可持续有重要意义。
4.1.3哈尼梯田传统农耕形成的水稻土与森林土微生物功能特征差异
哈尼梯田水稻土与森林土样品中不同菌群功能差异较大,不同的利用方式是产生物种差异的主要原因,而菌群差异导致土壤功能差异。同时,土壤作为一个非常复杂的生态系统,森林土与水稻土相比,森林土的异质化程度更高,水稻土进行均一化耕作后,其微生物结构功能特征的变异相对较小,而森林土壤由于在自然状态下存在较多变异从而进一步导致功能差异。在哈尼梯田传统耕作方式下,水稻土样品中所含菌群的功能主要有铁呼吸、硫化物呼吸等与元素循环有关的功能(图8),这与哈尼梯田水稻土中的核心菌群有着密不可分的联系。郭莹等[45]发现长期施用粪肥有助于提高土壤微生物的碳源利用能力,增强土壤有机质的转化周转能力;
罗希茜等[46]发现有机肥与化肥配合施用可以使微生物的碳源利用率明显增强;
拓晓骅等[47]在研究淹水培养的水稻地杆菌科微生物群落结构时,提出地杆菌科微生物对铁的生物地球化学循环具有重要影响。水稻土核心微生物如地杆菌、厌氧粘细菌不仅在全球碳循环中发挥了重要作用,还是许多沉积物中主要的铁还原微生物。这些微生物在修复重金属污染水体时发挥功能,如通过添加乙酸或其他有机质,可有效刺激地杆菌属微生物的大量富集,对维持水稻土壤环境有着十分重要的作用[42]。除施肥方式外,人为耕作的翻耕行为也会对土壤微生物有较大影响。人为耕作方式下的水稻土壤微生物的氨化作用增强,具有氨化作用的互营菌与森林土相比也有所增加。肖嫩群等[48]发现在翻耕方式下,土壤氨化作用以及氧化作用增强。对比自然状态下的森林土,其土壤中由于存在植物残体、落叶及其他含有木质素等难降解的有机质,故样品中菌群的功能主要涉及纤维素降解、木质素分解等。秦余等[49]在针阔叶混交林中采样分析得出森林土中优势菌种为嗜酸栖热菌、独活假丝酵母等有机质降解功能明显的菌群,这与我们的研究具有一致性。不同的微生物群落结构决定了哈尼梯田水稻土与森林土微生物功能上的较大差异。特别的,哈尼梯水稻土壤中微生物具有与元素循环有关的功能潜能。
4.2 结论
与森林土壤相比,传统农耕方式下的哈尼梯田水稻土壤微生物多样性显著增加。在门分类水平上,哈尼梯田水稻土的优势物种为绿弯菌,森林土的优势物种为放线菌。在属水平上两者差异更加显著,水稻土中嗜酸栖热菌的数量比森林土中显著减少,而厌氧粘细菌显著增加。地杆菌是哈尼梯田水稻土的关键指示微生物,具有生物修复等功能。慢生根瘤菌是森林土的关键指示微生物。通过CCA发现,总磷与水稻土壤微生物群落的相关性最高,淹水深度和pH与森林土壤微生物群落相关性最高。在哈尼梯田传统农耕方式下的水稻土中,微生物具有更高的铁呼吸、硫呼吸、好氧亚硝酸盐氧化功能潜力,与原始森林土相比,传统耕作方式显著重塑了土壤微生物群落结构及功能特征。
猜你喜欢 耕作群落哈尼 江垭库区鱼类群落组成和资源量评估当代水产(2022年8期)2022-09-20论丝竹玩友——群落生态视野下的乐人群体考察(下)中国音乐学(2022年2期)2022-08-10基于数字孪生的农业耕作监控系统农业技术与装备(2022年3期)2022-05-31首届哈尼梯田无人机摄影(像)大赛获奖作品云南画报(2021年4期)2021-07-22人教版生物必修3第4章第4节群落的演替教学设计学校教育研究(2020年7期)2020-04-09两伊握手环球时报(2019-03-12)2019-03-12特朗普放话要会鲁哈尼环球时报(2018-08-01)2018-08-01种群和群落及生态系统考题分析中学生理科应试(2017年6期)2017-09-27小麦种植不同耕作措施的对比江苏农业科学(2015年9期)2015-10-20