邱 妍,翁启杰,李 梅,陈升侃,3,周长品,李发根,甘四明*
(1.中国林业科学研究院热带林业研究所,热带林业研究国家林业和草原局重点实验室,广东 广州 510520;
2.南京林业大学,江苏 南京 210037;
3.广西壮族自治区林业科学研究院,中南速生材繁育国家林业和草原局重点实验室,广西优良用材林资源培育重点实验室,广西 南宁 530002)
林木生长既是对光照和气温等环境条件的适应性表现[1],也是用材树种育种的基本目标性状[2]。林木为多年生植物,其生长过程有自身的节律,并受环境条件的影响[1,3]。目前,已在多类树种中报道了生长性状的遗传力和遗传相关等遗传参数的研究,如桉属(Eucalyptus L"Hérit.)[4-23]和松属(Pinus Linn.)[24-25]树种,但多数研究仅分析了单个年份或早期的生长性状。因此,需要加强林木多年生生长的遗传分析,这有助于加深对林木生长动态遗传基础的理解,也将为早期选择提供有效指导。
尾叶桉(E.urophylla S.T.Blake)和细叶桉(E.tereticornis Smith)均为桉属双蒴盖亚属(Symphyomyrtus (Schauer) Brooker)树种[26],前者天然分布于东帝汶和印度尼西亚的一些岛屿(7°23"~10° S),后者分布于巴布亚新几内亚的南部至澳大利亚的维多利亚州南部沿海的狭长区域(6°~38° S)[27-28]。两树种亲缘关系较近,其杂种常用于遗传参数的估算[9-10,16,20]。尾叶桉×细叶桉杂种具有较好的速生、高产、抗风和耐寒潜力[19,29-30],已较广泛地用于华南地区桉树人工林的营建。
本研究对尾叶桉×细叶桉不完全析因交配产生的59个杂种组合13、16年生的树高、胸径和材积进行了狭义遗传力估算,并结合前期生长和材性[9]进行了遗传相关分析,以期进一步了解桉属树种生长动态的遗传规律,并为大径材良种培育的早期选择年龄的确定提供可靠依据。
1.1 参试材料与试验设计
参试材料为10株尾叶桉母本与10株细叶桉父本不完全析因交配产生的62个杂种组合,大田试验最终保存59个组合[9,20];
参试材料也包括10个母本的自由授粉家系。2003年7月在福建省漳州市长泰县岩溪林场美山工区(117°52′ E,24°46′ N)营建试验林,试验地概况见文献[20]。试验设计为随机完全区组,4株单行小区,4次重复,株行距为2 m×3 m。
1.2 性状测定
生长性状包括树高(H)、胸径(D)和材积(V)。利用Vertex III激光超声波测高测距仪(瑞典Haglof公司)测量13、16年生的树高(分别缩写为H13和H16),利用胸径尺测量13、16年生的胸径(D13和D16)。前期生长包括0.5、1.5、2.5、4、7.5、10年生的树高(依次缩写为H0.5、H1.5、H2.5、H4、H7.5、H10)和1.5、2.5、4、7.5、10年生的胸径(依次为D1.5、D2.5、D4、D7.5、D10)及1.5、2.5、4、7.5、10、13、16 年生的材积(依次为 V1.5、V2.5、V4、V7.5、V10、V13、V16),按文献[16]的公式计算:
V=H×D2/30 000
10年生材性的测定见文献[9]。基于之前建立的预测模型[10],利用近红外光谱仪MPA (德国Bruker)测定10年生木材基本密度(BD10)、纤维素含量(CC10)、半纤维素含量(HC10)、木质素含量(LC10)和紫丁香基木质素/愈创木基木质素比例(S/G10)。
1.3 统计分析


生长性状不同林龄间、生长与材性性状间的表型相关系数和加性遗传相关系数估算公式如下:
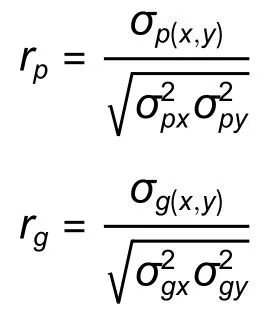
基于主成分回归进行16年生(因变量Y)与早期生长(树高、胸径和材积)的回归分析,模型如下:
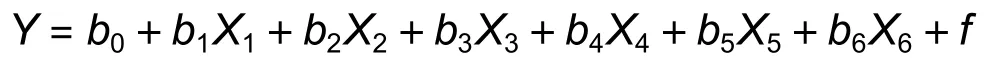
式中:b0为常数,b1~b6为回归系数,X1~X6依次为1.5、2.5、4、7.5、10、13年生生长经主成分变换后的自变量矩阵,f为误差。
2.1 尾叶桉×细叶桉生长表现
尾叶桉×细叶桉59个杂种组合13、16年生平均树高、胸径和材积均高于母本自由授粉家系(表1),表现出一定的杂种优势,尤其是13年生平均树高及13、16年生平均材积显著或极显著高于母本自由授粉家系(P≤0.05或0.001)。杂种组合的树高变异系数较小,但材积变异系数较大。
参照大径材(胸径≥28 cm)的标准[33],只16年生时1个杂种组合(UX-07×T2508,11株)的平均胸径达28.34 cm,13、16年生时分别有来自21和28个杂种组合的29和47株达到大径材的规格(表1),这为进一步培育大径材良种(包括无性系)提供了备选材料。

表1 尾叶桉×细叶桉杂种组合及母本自由授粉(OP)家系的生长及其变异系数(CV)Table 1 Growth and coefficient of variation (CV) of Eucalyptus urophylla×E.tereticornis crosses and maternal open-pollinated (OP) families
2.2 生长性状的遗传效应
表2表明:母本方差分量对13、16年生树高、胸径和材积均影响极显著(P≤0.001或0.01);
父本方差分量只对13、16年生材积影响极显著(P≤0.01),对树高和胸径均影响不显著(尤其是树高的方差分量接近0);
母本×父本互作的方差分量仅对13年生树高影响显著(P≤0.05)。这表明母本效应显著影响杂种组合的子代生长,而父本效应仅对材积性状影响显著,母本对子代生长的影响明显大于父本;
同时,母本×父本互作效应仅对某些年龄的树高有显著影响,且随着年龄增长呈减弱趋势。

表2 尾叶桉×细叶桉生长的母本、父本和母本×父本的方差分量及单株狭义遗传力和显性遗传效应Table 2 Female male and female×male variance components as well as single-tree narrow sense heritability and dominance ratio of phenotypic variancefor growth of E.urophylla×E.tereticornis crosses
2.3 性状间的表型相关和加性遗传相关
除1.5年生材积外,16年生树高、胸径和材积与之前年龄的相应生长性状的表型相关和加性遗传相关均达0.01或0.001的显著水平(图1),且加性遗传相关系数均大于对应的表型相关系数,这表明遗传相关是影响表型相关的重要因素。在2.5年生时加性遗传相关即较稳定,表明对16年生生长性状的早期选择可早至2.5年生时进行。

图1 16年生树高(H16),胸径(D16)和材积(V16)与之前年龄相应生长的表型相关和加性遗传相关Fig.1 Phenotypic and additive genetic correlations of 16-year-old height (H16),breast-high diameter (D16) and volume (V16) with earlier height,breast-high diameter and volume,respectively
13、16年生生长与10年生材性性状的表型相关系数和加性遗传相关系数见表3。无论是树高、胸径还是材积,13年生生长与材性的相关显著性与16年生类似(树高与木质素含量(LC10)的加性遗传相关除外)。13、16年生生长与10年生基本密度(BD10)均呈显著正相关(P≤0.01、0.05或0.001),而与LC10均呈显著负相关(P≤0.05或0.01,H16的rg除外),表明13、16年生时对生长的选择可间接提高木材基本密度,并降低木质素含量;
13、16年生生长与10年生紫丁香基木质素/愈创木基木质素比例(S/G10)均呈显著的表型正相关(P≤0.01或0.001),胸径和材积与纤维素含量(CC10)均呈显著的表型负相关(P≤0.05或0.01),但加性遗传相关均不显著,3个生长性状与半纤维素含量(HC10)的表型相关和加性遗传相关均不显著。
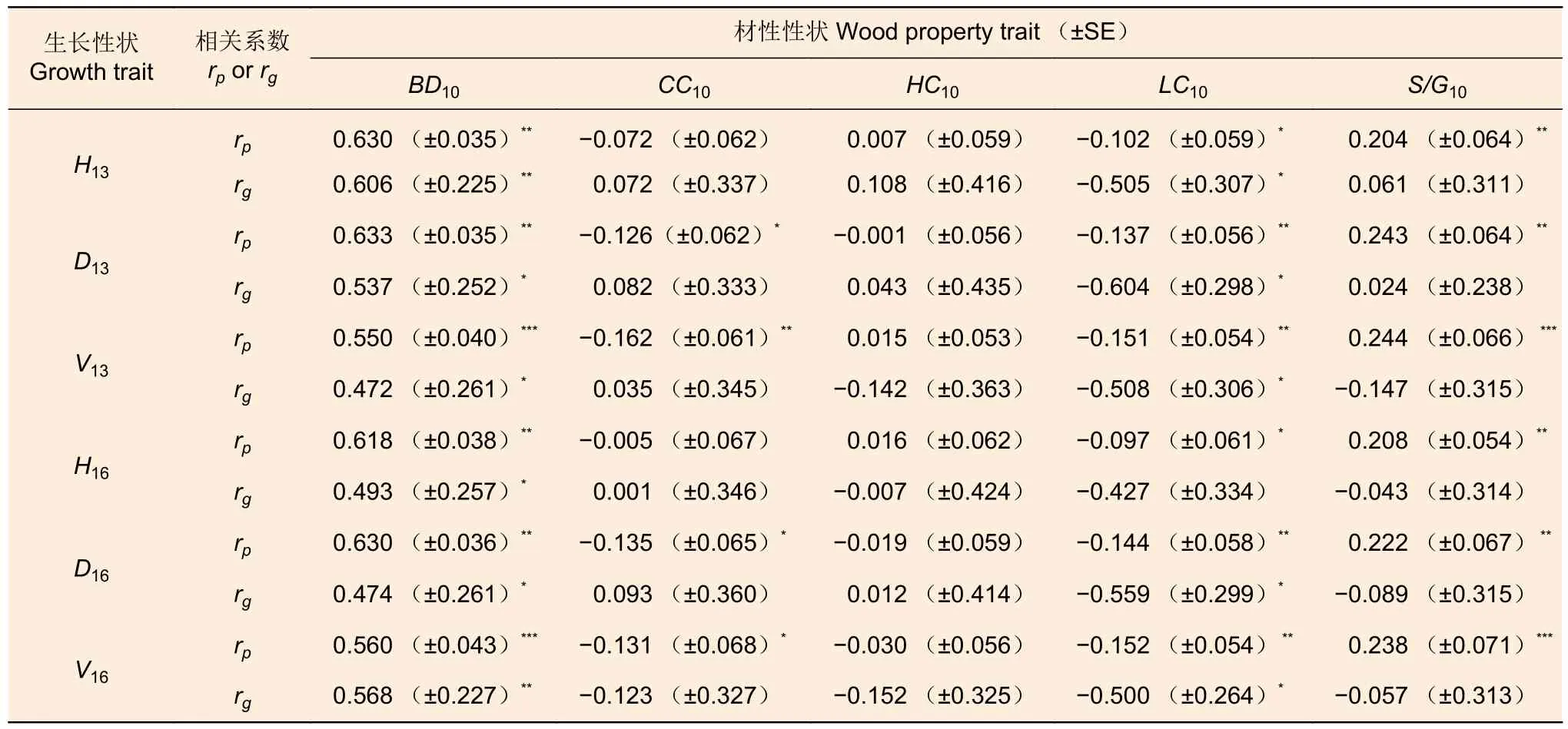
表3 13、16年生生长与10年生材性的表型相关(rp)和加性遗传相关(rg)Table 3 Phenotypic (rp) and additive genetic (rg) correlations of 13-and 16-year-old growth with 10-year-old wood properties
2.4 生长性状的回归分析
16年生与早期树高、胸径和材积回归分析的决定系数分别为0.842、0.942、0.960,第1主成分可分别解释83.0%、91.3%、93.7%的表型变异(表4),其中,2.5年生树高、胸径和材积可分别解释16年生时的73.9%、71.9%、66.2%的表型变异,表明2.5年时进行早期选择是可行的。
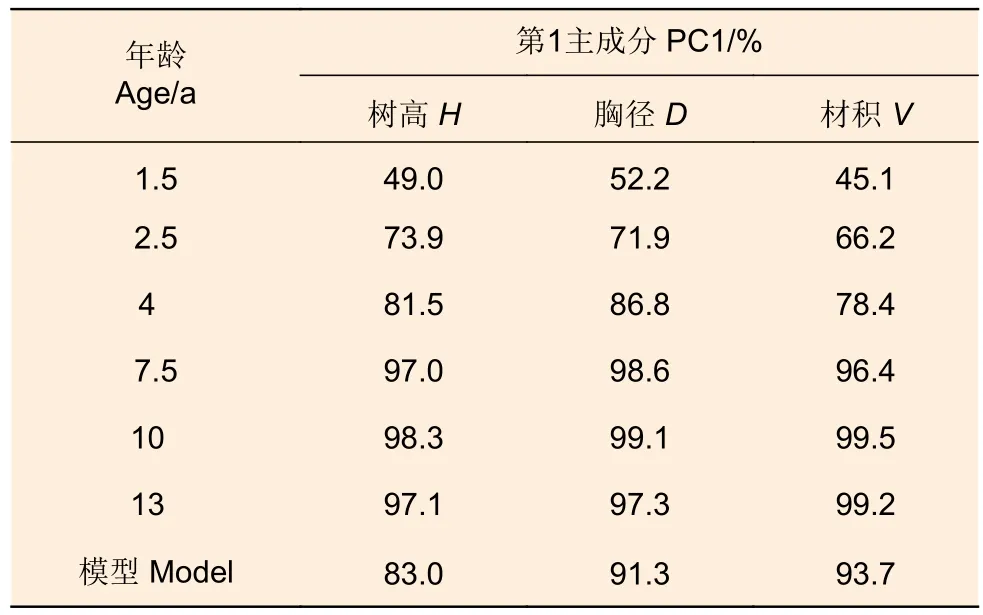
表4 16年生与早期生长回归分析的第1主成分Table 4 PC1 of principal components regression of 16-year-old growth with earlier growth traits
尾叶桉×细叶桉早期生长的杂种优势已有报道[20,29-30]。本研究中,杂种组合13、16年生的平均生长均好于母本自由授粉家系,表明杂种优势持续存在,其中,1个杂种组合16年生的胸径达到了大径材的规格,后续可利用其亲本建立种子园,生产(母本的)优良种子用于营林实践,从而实现杂种优势的遗传增益。尾叶桉×细叶桉易于无性快繁[34],杂种组合的优良单株经无性系测试后亦可大规模地快速推广。
尾叶桉母本的效应显著影响杂种13、16年生生长,这与7.5年生[20,30]和10年生[9]的研究结果一致,表明母本对尾叶桉×细叶桉整个生长期均存在显著的遗传效应。显著的母本效应也存在于尾叶桉×赤桉(E.camaldulensis Dehnh.)5年生[12]和8.3年生[22]生长性状中。相反,父本效应对13、16年生树高和胸径的影响不显著,这与10年生生长[9]的研究结果一致,但异于7.5年生生长[20,30]的研究结果,表明父本效应可能随着年龄增长而趋于减弱。另一方面,父本效应对材积的影响显著。可见,杂交育种中需要重视杂交力高的优良母本和父本的选择。此外,母本×父本互作效应对13、16年生生长(除H13外)的影响均不显著,这与7.5年生[20,30]和10年生树高、胸径[9]的研究结果相似,但与对1.5年生和2.5年生的显著影响[20]相反,表明母本×父本互作效应随年龄减弱,且程度可能比父本效应更大。当然,这并不能说明大径材良种培育中可以忽略母本×父本互作(特殊杂交力)的效应,因为,这些效应仍有一定程度的影响,尤其特定环境下或对生长之外的其他性状可能影响显著[9,16]。

早晚期生长的相关分析及主成分回归分析为尾叶桉×细叶桉早期选择提供了理论支持。类似地,蓝桉16年生与4年生胸径的加性遗传相关达0.95[5],23年生与3、11、17年生胸径的遗传相关分别为0.53、1.00、1.00[8],均表明3年生左右进行早期选择是可行的。采用主成分回归分析法估算了火炬松(P.taeda Linn.)[35]早期选择年龄,表明7年生胸径能够解释19年生胸径变异的88.13%。另一方面,尾叶桉×细叶桉晚期生长与木材基本密度和木质素含量的显著相关也为间接选择提供了可能。类似地,蓝桉16年生胸径与木质素含量呈显著的遗传负相关[6]、与基于Pilodyn检测的6年生木材密度呈显著正相关[5];
但本研究中一些性状间的相关与之前的报道有异,如尾叶桉14年生树高和胸径与木材基本密度呈显著的遗传负相关[7],蓝桉16年生胸径与纤维素含量和S/G比例均呈显著的遗传相关[6],这种差异可能是由于不同树种、地点和年龄引起的[11,16]。
尾叶桉×细叶桉13、16年生生长具有一定的杂种优势,具备大径材选育的潜力,这为下一步培育大径材良种(包括无性系)提供了备选材料。多年生生长性状持续受母本效应的显著影响,13、16年生时父本效应只对材积影响显著,杂交育种中需要重视优良母本和父本的选择。16年生与早期树高、胸径和材积遗传相关及回归分析均显示,在2.5年生时可解释16年生生长变异的66.2%~73.9%,表明尾叶桉×细叶桉大径材良种培育的早期选择可早至树龄2.5年时进行。晚期生长与10年生木材基本密度和木质素含量分别呈显著的正相关和负相关,对晚期生长的选择可间接提高木材基本密度,并降低木质素含量。
猜你喜欢 父本母本胸径 马尾松公益林胸径分布规律及冠幅影响因子分析湖南林业科技(2022年4期)2022-08-29甘肃祁连山森林资源连续清查中祁连圆柏前后期胸径关系的探究防护林科技(2021年3期)2021-09-12传统《元宵歌》本的复现、消失与走向人文杂志(2017年9期)2018-03-10杂交粳稻常优2号母本机插制种技术现代农业科技(2016年22期)2017-03-24父本在杂交谷子化学成分分布中的评价作用江苏农业科学(2015年5期)2015-10-20鹦鹉鱼品系对不同亲本和繁殖技术的响应科技资讯(2014年26期)2014-12-03用地径胸径回归分析法推算采伐木蓄积南方农业·下旬(2014年2期)2014-09-23玉米杂交制种中父本的管理技术现代农业科技(2009年1期)2009-04-09甜(辣)椒杂交制种育苗技术现代农业科技(2009年17期)2009-03-08